Эритроциты
Постоянное образование клеток крови, в том числе и эритроцитов, происходит у млекопитающих в стволовых клетках красного костного мозга. Полипотентные стволовые клетки производят клетки-предшественники, которые затем дифференцируются на клетки эритроидно-го, гранулоцитарного, мегакариоцитарного и агрануло-цитарного (моноцитарного и лимфоидного) рядов. Конечным результатом этого процесса является выход в кровеносную систему эритроцитов, лейкоцитов и тромбоцитов (рис. 5, б).
При различных патологических состояниях животного могут наблюдаться регенераторные (с достаточной функцией костного мозга) и гипопластические (включающие в себя гипорегенераторные и арегенераторные) изменения эритроцитов.
Эритроциты — наиболее многочисленная группа форменных элементов крови. Эритроциты млекопитающих — это безъядерные структуры, имеющие форму двояковогнутых дисков. Эта особенность формы эритроцитов отчетливо различима в мазках крови. Так, в мазках крови у эритроцитов различают бледную область в центре, окруженную более темным кольцом. Такая неравномерность окрашивания связана с двоякой вогнутостью эритроцита при горизонтальном положении его на предметном стекле: более тонкая центральная область окра-
Схема гемопоэза
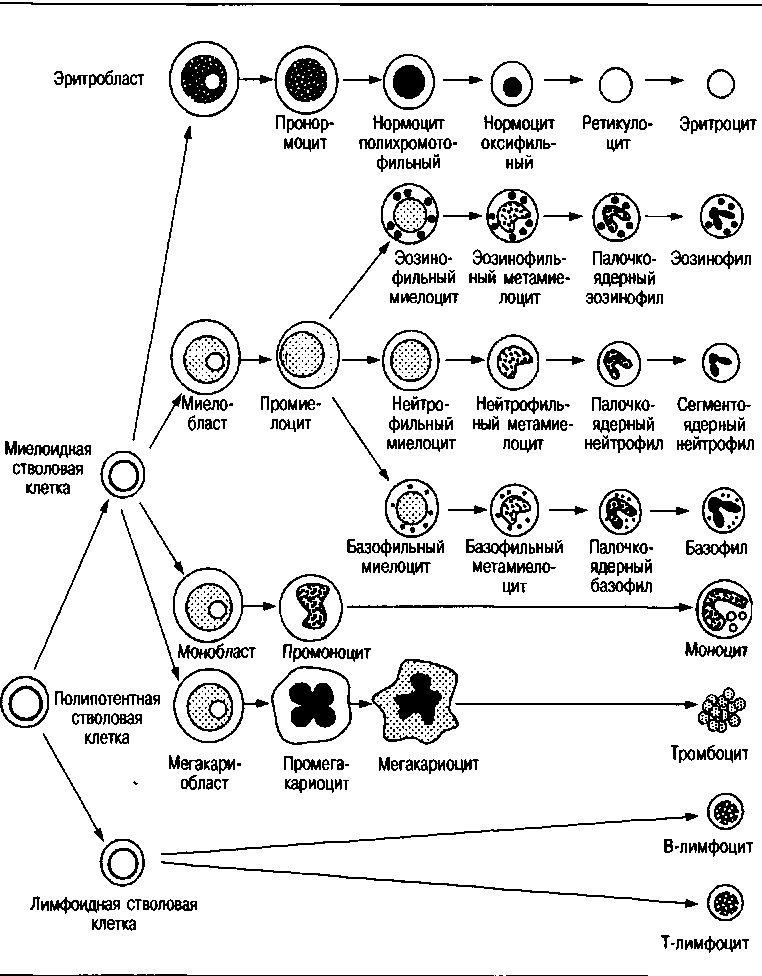
шивается бледнее, более толстая, периферическая — интенсивнее.
Центральная бледная зона легко различима в эритроцитах у собак, менее заметна —у кошек и лошадей (рис. 2, 3).
Двояковогнутая форма обеспечивает эритроцитам выполнение их основной функции — перенос газов, так как при такой форме поверхность диффузии увеличивается, а расстояние диффузии уменьшается. Кроме того, благодаря своей форме эритроциты обладают большей способностью к обратимой деформации при прохождении через узкие изогнутые капилляры.
В условиях нормы эти эритроциты никогда не оставляют кровеносную систему, исключение составляет красная пульпа селезенки, в которой имеются участки незамкнутости кровеносных сосудов.
Большая часть эритроцитов (66 %) имеют форму двояковогнутого диска. При этом возможны и другие формы эритроцитов: в виде шапочки — стоматоциты (18,5%), округлые с шипиками — эхиноциты (5,7%), округлые, гладкие — сфероциты (4,2 %) и другие. Изменение формы эритроцита по сравнению с самой распространенной называется пойкилоцитозом.
Диаметр эритроцита у собак 5—9 мкм (среднее — 7 мкм), кошки 4,9 мкм, лошади 4—7,5 (среднее — 5,6 мкм), 2,6 мкм толщиной по краям и 0,8 мкм — в центре. Средняя продолжительность жизни эритроцита различается у различных видов домашних животных. Так, у собак она составляет ПО—120 дней; у кошек — 66—79 дней; у лошадей — 140—150 дней. Размеры и количество эритроцитов у различных видов животных различаются, при этом имеется следующая закономерность: чем меньше эритроцит, тем большее их количество приходится на единицу объема крови. Собаки имеют наибольшие среди всех домашних животных эритроциты (d = 7,0 мкм), наименьшие у кошек (d = 4,3 мкм).
Изменение размеров эритроцитов называется анизоци-тозом. Это обычное явление для большинства видов животных.
У зрелых эритроцитов нет ядра и органелл. Цитоплазма заполнена гемоглобином, сложным белком, способным нестойко связываться как с кислородом, так и углекислым газом. Именно эти свойства гемоглобина обеспечивают основную функцию эритроцитов — газообмен.
По содержанию гемоглобина эритроциты неодинаковы. Их подразделяют на нормо-, гипо- и гиперхромные. Способность эритроцитов переносить кислород зависит от количества гемоглобина в них. Снижение как количества эритроцитов, так и количества гемоглобина в них приводит к нарушению окислительных процессов в организме и развитию гипоксии.
Гемоглобин (НЬ) в крови животного (в норме и патологии) присутствует в нескольких формах: оксиге-моглобин (НЬ02); дезоксигемоглобин, или восстановленный гемоглобин (НЬН); карбоксигемоглобин (НЬС02); сульфгемоглобин (БНЬ); нитрозогемогло-бин (НЬЫО); метгемоглобин (НЬМе1) или гемиглобин (Н|); цианметгемоглобин, или гемиглобинцианид (СЫтеШЬ).
В капиллярах легкого под влиянием высокого парциального давления в плазме крови кислород активно диффундирует в эритроциты, где, соединяясь с гемоглобином, образует оксигемоглобин. В тканях организма, где парциальное давление кислорода низкое, он отщепляется от оксигемоглобина. В результате чего возникает дезоксигемоглобин, к которому присоединяется выделяемый тканями углекислый газ (С02), в результате чего образуется кар-
боксигемоглобин. Большая же часть С02 поступает в кровь из тканей и оттуда — в эритроциты. В эритроцитах С02, соединяясь с водой, формирует угольную кислоту (Н2С03), которая диссоциирует на ионы. Затем в виде бикарбонатного иона (НС03~) выделяется обратно в плазму крови.
В капиллярах легких НС03~ из плазмы крови возвращаются в эритроциты, где вновь формируется Н2С03, которая распадается на Н20 и С02. Затем С02 выходит из эритроцитов в плазму крови, а далее — в просвет альвеол легких.
В норме в крови циркулирует до 1 % «юных» эритроцитов, называемых ретикулоцитами (рис. 4). В их цитоплазме, при окрашивании кризиловым фиолетовым либо метиленовым синим, выявляются сетевидные структуры — небольшое количество рибосом, обеспечивавших во время предшествующего созревания и диффе-ренцировки эритроцита синтез глобина. Ретикулоциты дозревают до эритроцитов в кровотоке за 24—30 часов. Повышенное количество ретикулоцитов в крови указывает на высокую потребность организма в кислороде, которая может быть следствием недавнего кровотечения.
Перенося кислород, эритроцит не потребляет его и не расходует на это энергию, так как энергию получают путем анаэробного гликолиза.
Примерно с 60-го дня жизни эритроцит начинает стареть и «изнашивается», при этом его пластичность уменьшается. На строение и функции эритроцитов большое влияние оказывает осмотическое давление плазмы крови. В гипотоническом растворе эритроциты набухают, и гемоглобин из них выходит. Гемолиз эритроцитов вызывают растворители жиров, например липолитиче-ские ферменты змеиного яда, плазма чужой крови и др.
В гипертоническом растворе вода уходит из эритроцитов, вследствие чего они сморщиваются.
Стареющие эритроциты элиминируются в основном в селезенке, частично в печени и костном мозге, где распознаются и фагоцитируются макрофагами. Сигналом для фагоцитирования макрофагами изношенных эритроцитов является изменение олигосахаридов их мембран и наличие на Рс-рецепторах плазмолеммы эритроцита комплексов «антиген — антитело».
По мере старения в эритроцитах вначале понижается, а затем нарушается гликолиз, вследствие чего их энергетический потенциал снижается. Одновременно с этим в стареющих эритроцитах нарушается цитоскелет, что приводит к изменению их формы. Из двояковогнутого диска вначале они приобретают строение стоматоцитов, затем — эхиноцитов, сфероцитов и других форм.
Параллельно с изменением цитоскелета при старении эритроцитов изменяются олигосахариды их плазмолеммы. На плазмолемме эритроцитов имеются Рс-рецепто-ры к иммуноглобулинам. На них фиксируются комплексы «антиген — антитело». Количество фиксированных иммунных комплексов в физиологических условиях возрастает постепенно, параллельно со старением эритроцитов. В случае же, когда в организме активно протекают иммунные реакции с формированием иммунных комплексов (например, при гнойно-воспалительных, аутоиммунных заболеваниях), Рс-рецепторы плазмолеммы эритроцитов становятся «забитыми» ими. «Нагруженные» иммунными комплексами еще не состарившиеся эритроциты макрофаги селезенки принимают за «состарившиеся» и фагоцитируют их, что приводит к понижению количества эритроцитов в крови, хотя эрит-ропоэз при этом не страдает.
В макрофагах селезенки, фагоцитировавших эритроциты, гемоглобин эритроцитов расщепляется на билирубин и гемосидерин (пигмент, содержащий железо).
Гемосидерин, превращаясь в трансферрин, поступает в макрофаги костного мозга, а из них — в развивающиеся эритроциты. Билирубин переносится в печень, где преобразуется в биливердин, входящий в состав желчи. В печени же накапливается обычно и некоторое количество гемосидерина. Его количество может доходить до значительных размеров в патологических условиях, когда происходит усиленный распад эритроцитов и гемоглобина. Образующийся при этом гемосидерин накапливается в значительных объемах в печени, в селезенке, в костном мозге и в лимфатических сосудах, обуславливая их гемосидероз.
Изучение мазка периферической крови было и до сих пор остается важной частью гематологического исследования, несмотря на широкое внедрение в клиническую практику автоматизированных анализаторов, которые, несомненно, гораздо более эффективны, чем ручные методы исследования, при определении средних величин и количественных гематологических характеристик. Тем не менее клетки крови изучаются врачом-лаборантом в мазках, что обусловлено малой надежностью, а зачастую и абсолютной непригодностью автоматизированного анализа для выявления редких аномалий форменных элементов крови (например, ядросодержащих эритроцитов, фрагментов эритроцитов).
Раньше количество эритроцитов в крови подсчитывали в счетной камере (камера Горяева), в настоящее время для этих целей широко используют гемоанализаторы, которые идентифицируют эритроциты по их объему.
Такой подсчет может давать погрешность в размере до 1 %. Однако в случаях значительного увеличения в крови других форменных элементов, по объему сопоставимых с эритроцитами (например, лимфоцитов), может быть ошибочно определено увеличение числа эритроцитов в крови. По мнению I. Б. Веввтап (1980), такие артефакты встречаются редко, но тем не менее о них следует помнить.
⇐ФОРМЕННЫЕ ЭЛЕМЕНТЫ КРОВИ || Оглавление || Изменения количества эритроцитов в крови⇒